
Abstract
Threats to coastal wetlands, including sea-level rise and subsidence, led the National Wildlife Refuge (NWR) System to protect over 500,000 hectares of coastal wetlands during the twentieth century, with approximately 20% occurring in the South Atlantic geography. This effort has involved systematic long-term monitoring of changes in marsh elevation using surface elevation tables and marker horizons at 20 sites across 19 NWRs in the southeastern coastal USA. From 2012 to 2021, the rates of change in surface elevation (−9.3 to 7.1 mm/year), accretion (−0.3 to 17.5 mm/year), and net vertical elevation change (−14.3 to 3.1 mm/year) were highly variable among monitoring sites and varied with coastal wetland type (oligohaline marsh, salt marsh, pocosin, or forested wetland), land surface elevation, and estuarine salinity and geomorphology (i.e., tidally influenced or embayed). Of 20 sites included in our study, only six were gaining elevation at a rate that was equal to or greater than the long-term rates of sea-level rise and therefore considered resilient. Only Waccamaw and Currituck NWRs, both located in oligohaline marshes, were gaining elevation at a rate that exceeded sea-level rise by 1 mm/year. These results support the mounting evidence that many coastal wetlands, particularly in the South Atlantic geography of the USA, will undergo ecological transformations in the next several decades. The NWR System and other coastal management entities will need to use strategic decision-making frameworks to identify management actions that can mitigate the loss of coastal wetlands to support the conservation of coastal wetland–dependent and obligate species.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Coastal wetland ecosystems absorb the energy of storms, improve water quality in bays and estuaries, supply nutrients to marine food webs, provide critical habitat for birds and wildlife, and act as a nursery for many commercially important species of fish and shellfish (Daily et al. 1997; Barbier et al. 2011; Costanza et al. 2014; ACJV 2019). Wildlife species that depend on coastal wetlands, such as secretive marsh birds, fish, and waterfowl, are some of the U.S. Fish and Wildlife Service’s (USFWS) highest conservation priorities (ACJV 2019).
The future of our coastal wetlands and the wildlife that depend on these habitats is uncertain. Two recognized threats to coastal wetland stability are the projected increasing rates of relative sea-level rise (SLR) and subsidence, a subsurface process that often results in the loss or sinking of land surface elevation (Kirwan and Megonigal 2013; Kirwan et al. 2016; Osland et al. 2022a; Saintilan et al. 2022; Sweet et al. 2022). In the USA, recent climate models suggest an increase of SLR along the Mid-Atlantic coast that is greater than the global average due to glacial isostatic adjustment, which is the ongoing movement of land from glacial melting and groundwater depletion (Sallenger et al. 2012; Karegar et al. 2016; Krasting et al. 2016; Saba et al. 2016; Sweet et al. 2017, 2022).
The uncertainty of the future of our coastal wetlands motivated the USFWS National Wildlife Refuge (NWR) System to undertake systematic long-term monitoring of coastal wetlands on NWRs across the South Atlantic geography to estimate vertical resilience to SLR (Covington 2020). Over 500,000 of the 40 million hectares of coastal wetlands nationwide (approximately 1.25% of all coastal wetlands nationwide) are protected by NWRs, and approximately one-fifth (100,000 hectares) of all coastal estuarine and freshwater wetlands of NWRs are located in the South Atlantic geography (NOAA 2016).
As part of a national effort, the NWR System in the southeastern USA designed and implemented a Coastal Wetland Elevation Monitoring (CWEM) program to monitor the potential resiliency of coastal wetlands on NWRs to SLR across the South Atlantic geography (Moorman and Rankin 2020). Here, we define coastal wetland resiliency as coastal wetlands with positive net vertical elevation change, i.e., the wetland gained elevation at a rate equal to or greater than the published long-term rate of SLR from NOAA during the period of 2012–2021 (Kamrath et al. 2019). Some types of coastal wetlands have demonstrated resilience through positive biophysical feedback loops that occur between wetlands and rising water levels (Vogel et al. 1996; Kirwan et al. 2016). Within the southeast Atlantic, the Marsh Equilibrium Model from Morris et al. (2002) suggests that high sediment loading promotes the long-term stability of coastal wetlands as long as sea-level rise remains below 12 mm/year.
Coastal wetlands occur in a transgressive landscape where transformations between ecosystems are based on elevation and salinity gradients, with salt marshes occurring on the seaward end of this gradient (Osland et al. 2022a). Ecological transformation can occur if enough pressures act on the ecosystem such that a new state is achieved. SLR and subsidence will likely result in ecological transformations in coastal wetlands. For instance, current salt marshes may be inundated by rising seas and transgress or move upslope and upriver into other wetland systems (Osland et al. 2022b). One measurable variable that can help predict coastal wetland vulnerability to ecological transformation is net vertical elevation change, which can be predicted by subtracting estimated rates of SLR from rates of coastal wetland surface elevation change (Craft et al. 2009; Kirwan et al. 2010; Stagg et al. 2016).
The ongoing goal of CWEM is to estimate rates of surface elevation change, accretion, and net vertical elevation change for priority coastal wetland habitats on NWRs across the South Atlantic geography. This effort has improved our understanding of coastal wetland resilience to SLR and the processes contributing to the resilience, or lack thereof, at each site and by coastal wetland type. Results from these monitoring efforts will also help managers make ecologically informed decisions with respect to conservation and management, such as if restoration or transformative actions should be considered or continued. Here, we examine four questions relevant to coastal wetland dynamics and management: (1) What are the rates of elevation change measured on each refuge within the CWEM network; (2) how do surface elevation change rates compare to SLR rates; (3) do trends in elevation change vary among habitat types; and (4) what roles do sub-surface processes and surface accretion play in surface elevation change in each of these habitat types?
Methods
Study Area
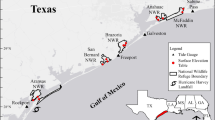
Oligohaline marshes, salt marshes, forested wetlands, and pocosins were initially selected as the priority habitats to be monitored on 18 coastal NWRs in the South Atlantic geography (Fig. 1). The South Atlantic geography was defined by the South Atlantic Landscape Conservation Cooperative and spans watersheds along the Atlantic coast from the Albemarle-Pamlico Sound in North Carolina and Virginia to part of the St. John River watershed in Florida (Fig. 1). It also includes the Apalachicola-Chattahoochee-Flint River basin on the pan handle of Florida. The focal region has a temperate climate with a mean annual precipitation and temperature range from 112 cm and 15 °C, respectively in Danville, Virginia to 147 cm and 20 °C, respectively, in Tallahassee, FL (Pickens et al. 2017).

Surface elevation table (SET) monitoring site locations in coastal wetlands along South Atlantic geography (gray polygon) within the USA. Four wetland habitat types are designated by colors showing oligohaline (yellow), pocosin (purple), salt marsh (green), and forested (blue). The locations of corresponding NOAA tide gauges are labeled as letters A-I and indicated as light blue triangles
We established CWEM monitoring sites (N = 20) in 2012 within the previously defined priority habitats using a spatially balanced random sampling design that selected a 0.5 ha unit (Moorman and Rankin 2020). These selected sites were accessible, had uniform vegetation cover with little disturbance, and were located at least 25 m away from water bodies and human structures (Moorman and Rankin 2020). Within each randomly selected site, three stations (experimental replicates) were established where marker horizon and surface elevation table (SET) measurements were made. In 2016, two additional sites located at Cape Romain NWR that had been sampled following the same protocol since 2010 were incorporated into the project. Ultimately, this monitoring effort included 19 NWRs, 22 sites, and 65 SET benchmarks in four states: North Carolina, South Carolina, Georgia, and Florida (Fig. 1). From 2012 to 2022, all sites were sampled following the USFWS regional protocol framework (Moorman and Rankin 2020), which is based on the protocol of Lynch et al. (2015). We measured plots quarterly for the first 2 years, semi-annually for the third year, and then annually thereafter (Moorman and Rankin 2020). Not all sites were measured during every sample period due to logistical and staffing constraints; only sites with a minimum of 5 years of data over the 10-year time period were used in our regional analysis (Fig. S1).
Sites were distributed along a gradient of coastal wetland types from the lowlands to the uplands: salt marsh (N = 13), oligohaline marsh (N = 6), forested wetland (N = 1), and pocosin (N = 2) and can be further classified as one of eight of NatureServe’s Ecological systems (Comer et al. 2003; Table 1, Fig. 1). Salt marsh sites included either embayed brackish marshes (N = 4) regularly flooded by storm surge and dominated by Juncus roemerianus in North Carolina, or tidal salt marshes with semidiurnal flooding dominated by Sporobolus alterniflora in South Carolina and Georgia (N = 7) or Juncus roemerianus in north Florida (N = 2). Oligohaline sites included both freshwater (N = 2) and oligohaline marshes (N = 4) with salinities generally less than five parts per thousand. Embayed oligohaline marshes in northeastern North Carolina were generally influenced by wind and tides and were dominated by Juncus roemerianus (N = 2). Tidally influenced oligohaline marshes of South Carolina, Georgia, and Florida were flooded daily and received additional pulses of water from storm surges and upstream flooding (Table S1). These oligohaline systems were dominated by Zizaniopsis millacea (Savannah and Ace Basin NWRs) or Cladium mariscus spp. jamaicense (Lower Suwannee NWR), and the freshwater site at Waccamaw NWR was dominated by Zizania aquatica. The forested wetland site at Roanoke River NWR was a blackwater swamp forest (Taxodium distichum—Nyssa aquatica—Nyssa biflora / Fraxinus caroliniana / Itea virginica) association that was flooded for extended durations following heavy rainfalls. Pocosin sites occurred on peat domes and were found in low-lying pond pine forest at Alligator River NWR and within scrub-shrub pocosin at Pocosin Lakes NWR. Pocosin sites were poorly drained but were upslope from tidal influences (Boyle et al. 2015; Cook et al. 2016).
Surface Elevation Monitoring
At each site, we installed three replicate SET benchmarks (i.e., stations) within the randomly selected 0.5-hectare sampling unit. Each benchmark was contained within a 3-m2 CWEM plot that was not disturbed by humans when sampling the site. A leveled SET arm was attached to the benchmark, and nine pins were placed in the arm that provide measurements of surface elevation (Fig. 2). These pin measurements were made in the same locations through time to estimate the rate of elevation change. Although 36 individual pin measurements (i.e., nine measurements in four cardinal directions) of elevation change were made around each SET station, they all were measuring the one station (the sampling unit) and are not considered independent replicates. However, replicates were represented by the three CWEM plots each containing a SET station within each site. More detailed information about the sampling protocol can be found in Moorman and Rankin (2020) and Lynch et al. (2015).

Surface elevation table (SET) monitoring apparatus. The level SET arm extends above the benchmark and nine equally spaced pins are placed through holes in the arm. The height of the pins above the SET arm are measured for long-term trends in elevation. Photo courtesy of Michelle Moorman, USFWS
We converted relative elevation measures to true surface elevation using the benchmark elevations measured at each SET station (Moorman et al. 2019), vertical offset from each SET apparatus, individual pin lengths, and pin height measurements using the following equation from Cain and Hensel (2018):
The true surface elevation (mm) for 2013 was computed for all sites to provide an estimate of the wetland elevation surface in the North American Vertical Datum (NAVD88). An estimate of a site’s surface elevation was then calculated for each subsequent sampling event.
We converted SET measurements of wetland surface elevation (mm) to the rate of change in surface elevation (mm/year) from the initial measurement at a time (t0), following methods described in Lynch et al. (2015). To calculate mean and variance estimates, slopes of regression models for each pin were calculated and then averaged for the four SET arm positions, the SET station, and the site (Lynch et al. 2015). Linear regression models were used to evaluate SET trends. We then computed an average rate of elevation change at the regional scale for the priority habitat types.
Benchmark Elevations
We surveyed SET benchmarks from 2015 to 2019 with Global Positioning System (Trimble 5700/5800 GPS Receiver, Westminster, CO, USA) equipment, which provided elevation measurements to 5-mm accuracy (Moorman et al. 2019). Benchmark elevation was assumed to be static to allow us to back-calculate the starting elevation of each station in 2013. GPS data were then processed using Online Positioning User Service (OPUS) Projects (Moorman et al. 2019). During OPUS sessions, a benchmark elevation was measured relative to the North American Vertical Datum of 1988 (NAVD88) using two consecutive static surveys of at least 2-h duration and post processed using Geoid 12a (Moorman et al. 2019). The estimated root mean squared error from OPUS sessions ranged from 13 to 304 mm. With these calculations, it should be noted that determining accurate elevations (sub-centimeter resolution) using GPS survey techniques was very difficult to achieve, and replicate surveys indicate that there is more error in the OPUS Projects network than previously reported. As such, benchmark elevations may have confidence intervals greater than 10–20 mm (Moorman et al. 2019).
Net Vertical Elevation Change
For reproducibility purposes, relative SLR rates as of 2021 were obtained from nearby National Oceanic and Atmospheric Association (NOAA) tide gauges (NOAA 2022; Table 2). Rates were calculated across all published NOAA tide gauge data, which date back to when the station was established (ranging from 1897 to 1978; Table S2). Considering SLR has been known to be accelerating over the past decades, our calculated rates of SLR were likely conservative (Dangendorf et al. 2019; Yin 2023). We compared these rates of relative SLR based on NOAA tide gauge data to rates of net vertical elevation change at each SET site. All NOAA tide gauges were located between 17 and 91 km from the closest corresponding SET site, which was calculated using the shortest geodesic distance between two points on an WGS84 ellipsoid (Table S2). We recognize that these SLR rates, due to their geographic location, may be different from the actual site.
Marsh surface elevation was compared with mean sea level to determine the elevation capital of a site (i.e., vertical distance between marsh surface and mean sea level). In addition to elevation capital, the net vertical elevation change, i.e., the difference between SET and SLR trends (mm/year), was computed to investigate whether coastal wetlands were keeping pace with SLR. Positive values of net vertical elevation change indicate a site was gaining elevation at a rate greater than the rate of SLR and could be considered resilient. Negative values of net vertical elevation change indicated a site was gaining elevation at a rate less than the rate of SLR. In other words, even if a site was gaining elevation, if it was doing so at a rate slower than SLR, it was at an elevation deficit relative to the long-term rate of SLR.
To test for differences in the relationship between SET to SLR elevation change among coastal wetland types, we fit a generalized mixed-effects linear model using the “lmerTest” package in program R (version 3.1–3; Kuznetsova et al. 2017). We included the net vertical elevation change as the response variable and coastal wetland habitat type as the fixed-effect predictor variable, with the site as a random effect. Where significant differences among habitat types were detected, we tested for pair-wise differences with Tukey’s post-hoc test using the “emmeans” package (version 1.7.5; Lenth 2022).
Accretion Monitoring
To measure accretion, we established three marker horizon plots within each of the three CWEM stations at each site, for a total of nine marker horizon plots per site. To establish marker horizon plots, feldspar layers were deployed following methods in Moorman et al. (2019) which follows methods from Lynch et al. (2015). Each time the SET benchmark was measured, up to three soil cores were taken from each of the three independently maintained marker horizon plots around that benchmark until the feldspar marker was retrieved in the core. If a marker was found, three measurements of soil depth (mm) above the feldspar layer were recorded for each core. While the McCauley corer was primarily used at the beginning of sampling, some sites transitioned to cryogenic coring due to difficulties obtaining a solid core using the original method. The coring method is documented within the USFWS SET database (USFWS 2022). If the feldspar layer was never recovered, no measurement was obtained (N = 7 sites; Fig. S1). The marker horizon plots needed to be re-laid after the marker had degraded to a point that it was no longer detectable, which usually occurred in 2-to-3-year intervals (Lynch et al. 2015; Moorman and Rankin 2020). Marker horizon plots were re-laid in a location that was distinct from the previous one, except at Cedar Island NWR where the plot was accidentally re-laid over the previous plot.
Marker horizon data were analyzed following methods described in Lynch et al. (2015) by first averaging measured distances (mm) from the top of soil core samples to the marker horizon to get plot-level means. Annual linear accretion trends (mm/year) and 95% confidence intervals for the three marker horizon plots were computed and then averaged together to estimate a mean accretion trend for each CWEM station and for each site. An estimate of subsidence was obtained by subtracting the SET trend from the accretion trend. Positive estimated subsidence values indicated that subsidence was the primary contributor to the overall SET trend, while negative estimated subsidence values indicated that belowground expansion was the primary contributor to the overall SET trend (Lynch et al. 2015). These data could then be used to make inferences about above and belowground processes driving SET trends and vertical elevation change at each sampling site among coastal wetland types. All data used within this manuscript are available in a public database and analyzed in program R (version 4.2.0; Ladin and Moorman 2021; R Core Team 2022; USFWS 2022).
Results
Trends and associated uncertainty could be calculated for 20 sites for surface elevation, and 13 sites for accretion. The surface elevation of SET sites was measured for 6–9 years during the period of 2012–2022 (Table 2; Fig. S1). We estimated the mean annual surface elevation trend, associated standard error, and 95% confidence intervals and compiled NOAA relative SLR trends with 95% confidence intervals for 20 CWEM sites (Table 2, Fig. 3). SET elevation trends were negative at the two pocosin sites at Alligator River NWR and Pocosin Lakes NWR. Mean SET trends did not differ from zero at three of the 13 salt marsh sites: Cape Romain — Horsehead Key, Cape Romain — Raccoon Key, and Blackbeard Island NWRs. Mean SET trends at all other sites were positive and ranged between 0.6 mm/year at St. Marks NWR and 7.1 mm/year at Waccamaw NWR (Table 2, Fig. 3).

Surface elevation trends (mm/year) across sites in the Coastal Wetland Elevation Monitoring network. Boxes show median (colored bars) and interquartile ranges with whiskers showing 1.5 times the interquartile range. Site colors denote wetland type: oligohaline (yellow), salt marsh (green), forested (blue), pocosin (purple). The range of sea-level rise rates is shown as a gray ribbon
The mean surface elevation (NAVD88) for each site in 2013 varied from −11 mm at Swanquarter NWR to 3164 mm at Pocosin Lakes NWR. Sites with low elevation capital at the start of the study (i.e., sites with less than 300 mm of elevation capital; Ganju, personal communication; Ganju et al. 2023) included the salt marsh sites at Alligator River, Swanquarter, Pea Island, and Cedar Island NWRs, the oligohaline marsh sites at Currituck and Mackay Island NWRs, and the forested wetland site at Roanoke River NWR (Fig. 4, Table 2). All sites were in the embayed estuary of the Albemarle-Pamlico sound of North Carolina. By the end of the study, the wetland surface elevation of all salt marsh sites with low elevation capital except the site at Cedar Island was equal to or below the estimated elevation of mean sea level suggesting these sites are likely experiencing present-day ecological transformations. Conversely, both oligohaline sites maintained their elevation capital, although minimal throughout the study (Fig. 4).

Site-level benchmark-adjusted surface elevation (mm) over time (years) in comparison with mean yearly sea levels from the nearest NOAA tide gauge (mm). Simple linear models with ± 95% confidence interval bands are overlaid with benchmark-adjusted surface elevation data showing oligohaline (yellow), salt marsh (green), forested (blue), pocosin (purple) wetland types, and sea level (blue)
Long-term SLR rates spanned a gradient from 2.2 to 5.1 mm/year across the region of study. Net vertical elevation change, i.e., the difference between surface elevation gains and sea level rise, was computed for all sites and ranged from −4.3 to 3.1 mm/year (Table 2). Six of the 20 CWEM sites gained elevation at a rate greater than or equal to the long-term rates of relative SLR, indicating these sites had positive net vertical elevation change and could be considered resilient. These sites included three oligohaline marshes at Savannah, Waccamaw, and Currituck NWRs and three salt marsh sites at Alligator River — Salt Marsh, Swanquarter, and Cedar Island NWRs. The sites at Waccamaw and Currituck had net vertical elevation change that was more than double all other resilient sites.
We found differences in mean net vertical elevation change among coastal wetland types (F-value = 47.8, numerator df = 3, denominator df = 15.9, P < 0.0001), as well as pair-wise differences in mean net vertical elevation change using Tukey’s post-hoc test between pocosin habitats and each of the other coastal wetland types (−14.5 ± 1.1 mm/year, P < 0.0001 in all cases). Oligohaline marsh surface net vertical elevation change (0.6 ± 0.7 mm/year) was marginally greater than salt marshes (−1.7 ± 0.5 mm/year, P = 0.058) and similar to forested wetland trends (−1.3 ± 1.6 mm/year, P = 0.64). Of the four coastal wetland types, only oligohaline wetlands within our study had a mean surface elevation trend greater than the corresponding rate of SLR (Fig. 5).

Net vertical elevation change (i.e., difference between the rate of surface elevation change and sea-level rise; mm/year) where positive values indicate the rate of surface elevation change is increasing faster than long-term rates of sea-level rise, and conversely, negative values indicate the rate of sea-level rise is increasing faster than surface elevation change. Box-and-whisker plots show median and inter-quartile ranges with station-level means (N = 59) overlaid. Whiskers indicate 1.5 times the interquartile range. Coastal wetland types are shown for oligohaline (yellow; squares are embayed sites and triangles denote tidal), salt marsh (green; squares are embayed sites, and triangles denote tidal), forested (blue), and pocosin (purple), and pair-wise significant differences are shown by letters A–C
We analyzed data from marker horizon plots at 13 sites (Table 3; Fig S1). Seven sites were excluded from the analysis because they either contained data that were likely measured incorrectly, or the feldspar plate was never recovered. Accretion rates at the sites included in our study were highly variable, ranging from −0.3 to 17.5 mm/year (Table 3). Estimates of shallow subsidence ranged from −3.7 to 15.0 mm/year (Table 3).
Discussion
Aside from the pocosin sites, most sites within our study gained or maintained surface elevation. When comparing surface elevation trends with rates of SLR, the net vertical changes at most sites were either negative or neutral suggesting that these coastal wetland systems are not resilient to SLR and will be unable to keep pace with long-term, short-term, and projected future rates of SLR. Siegert et al. (2020) recently suggested the Intergovernmental Panel on Climate Change’s projected increase in SLR of 0.61–1.10 m by 2100 (if global temperatures increase by 4 °C) may be conservative if the mass lost from glacial ice sheets was underestimated. As SLR occurs, we can expect increased flooding and inundation of our coastal wetland habitats. For USFWS, management strategies aimed at increasing the elevation capital of any site could increase the resiliency of the habitat and resist the transformation of the wetland in the short term as new habitats and migration corridors are developed.
The rate of surface elevation change in our study differed among coastal wetland types and estuarine geomorphology, i.e., whether the system was tidally influenced or embayed. Oligohaline marsh sites gained more elevation and exhibited higher net vertical elevation change in comparison to the other coastal wetland types (Fig. 5). These rates are comparable to those measured in other oligohaline and freshwater systems in the South Atlantic geography (Stagg et al. 2016). Surface elevation change can also be differentially affected by aboveground processes such as sediment erosion and deposition and belowground processes such as soil compaction and plant dynamics (Cahoon et al. 1998; 2021). Estimates of the rate of belowground and aboveground processes from accretion data varied by site and helped parse out the surface and subsurface processes that were affecting surface elevation dynamics. At sites where both the SET trend and estimated subsidence were positive, we posit that shallow subsidence was occurring and that accretion was driving the positive SET trends observed. We observed this phenomenon at Waccamaw, Savannah, ACE Basin, Alligator River-Salt Marsh, Swanquarter, Pea Island, and St. Marks NWRs. If the SET trend was positive and estimated subsidence was negative, we posit that shallow expansion was occurring and driving the positive SET trends. We observed this result at Currituck, Mackay Island, Cedar Island, and Roanoke River NWRs. If the SET trend was negative and the estimated subsidence was positive, we posit that subsidence was driving the negative SET trends. We only observed this result at the two pocosin sites at Alligator River-Pocosin and Pocosin Lakes NWRs (Table 3, Fig. 5). Sites undergoing submergence during our study period had negative or neutral vertical elevation change and little elevation capital.
Potential Processes Driving Vertical Migration in Tidally Influenced Oligohaline and Salt Marshes
At the tidally influenced oligohaline marshes of Waccamaw, Savannah, and ACE Basin NWRs, accretion (17.5, 8.9, and 6.9 mm/year, respectively; Table 3) was the main contributing factor to positive SET trends. High rates of accretion were also found in previous studies at Savannah NWR, Waccamaw NWR, and at a tidally influenced freshwater marsh in Georgia (20, 12, and 14 mm/year, respectively; Craft 2007; Stagg et al. 2016). Results from a study of wetlands along the Chesapeake Bay in the Mid-Atlantic region, just north of our study area, suggested inundation positively contributed to accretion in that system through sedimentation. This may help explain a mechanism for increased accretion rates in the tidally influenced marshes that were flooded daily (Palinkas and Engelhardt 2019). Similar to Saintilan et al. (2022), we observed that as accretion rates increased, so did the rate of subsidence in tidally influenced oligohaline systems. For instance, the Waccamaw site had the highest accretion rates, averaging 17.5 mm/year, but also had high estimated rates of subsidence, averaging 10.4 mm/year (Table 3, Fig. 6).

Mean marsh elevation trend (mm/year) for 13 U.S. Fish and Wildlife surface elevation table monitoring sites. Time periods for these data are shown in Table 3. Mean surface elevation trends and standard error are shown as red dots with error bars. Contributions of accretion as determined by the marker horizon trend (shown by the white bar graph) and subsurface processes (i.e., accretion trend subtracted by the SET trend; shown by the gray bar graph). Sites are grouped by habitat type
Conversely, accretion was minimal at our tidally influenced salt marsh site located on the upland edge of the marsh at St. Marks NWR in FL. Other studies of salt marshes in South Carolina and Georgia have found accretion rates to average 2.1 mm/year which is significantly less than the measured rate of accretion in the tidally influenced oligohaline marshes (11.1 mm/year; Crotty et al. 2020). Lower rates of accretion may provide the mechanism for reduced rates of elevation gains in tidally influenced salt marshes, suggesting there may be opportunities to enhance accretion in these marshes to increase the long-term resilience of these systems, particularly if elevation capital is high (Langston et al. 2021).
Potential Processes Driving Vertical Migration in Embayed Oligohaline and Salt Marshes
The rate of net vertical elevation change within the embayed oligohaline systems at Currituck and Mackay Island NWRs was similar to the tidally influenced oligohaline systems, but the rate of accretion was less than the rate of surface elevation change. This suggests that belowground expansion, rather than accretion, is contributing to the net change in surface elevation at these embayed oligohaline sites. We found similar results in the embayed salt marsh systems of North Carolina, where accretion was also minimal. The geomorphology of the embayed North Carolina salt marsh and oligohaline sites was unique in that they were microtidal, wind-driven systems, which are conditions that have been found to slow the rate of accretion (Lagomasino et al. 2013).
Our findings support the results from other studies that have measured low accretion rates of 0.9 to 2.4 mm/year in the embayed Albemarle-Pamlico system (Craft 2007; Lagomasino et al. 2013; Gundersen et al. 2021). Our estimates of accretion and subsidence suggest that belowground biomass, potentially because of enhanced vegetation growth, was augmenting elevation gains. There are several plausible reasons why vegetation growth was enhanced in embayed systems that need further investigation. Reed (2002) noted that landscape position and the starting elevation of the land surface are key factors in vertical elevation gains. As described previously, the embayed marsh sites in North Carolina had the lowest elevation capital of the sites within the study and simultaneously the highest rates of SLR (Table 2). Other studies have found that vegetation growth rates of salt marshes were significantly enhanced in low elevation zones and during years of higher sea levels (Morris et al. 2002; Silvestri and Marani 2004; Wasson et al. 2019) and that tidal amplitude can correspond to an energy subsidy that enhances the productivity of plants (Odum et al. 1995).
In addition to having low elevation capital, another plausible explanation is that many of the marshes within the embayed Albemarle-Pamlico system have been burned either through the application of prescribed fire or from lightning strikes. For instance, Mackay Island NWR, a site with prescribed fire at 5-year intervals (i.e., 2005, 2010, 2015, 2019, Mike Hoff, personal communication), exhibited shallow expansion at approximately 2.5 mm/year. In comparison, Swanquarter NWR, a nearby site located in a designated wilderness area and thus prohibited from applying active management techniques such as prescribed burns, exhibited shallow subsidence at 2.1 mm/year — the highest rate among the embayed marsh sites of the Albemarle-Pamlico system. Controlled burning of marshes has been shown to increase belowground biomass due to either increased root production or decreased soil decomposition processes and drive increases in elevation (Cahoon et al. 2004; McKee and Grace 2012). Collectively, these results suggest additional work should be undertaken to better understand the role of prescribed burns as a management strategy to increase belowground biomass and increase resiliency in coastal wetlands (Moon et al. 2022).
Potential Processes Driving Vertical Migration in Forested Wetlands and Pocosins
There were three non-marsh sites in our study that occurred in forested wetlands and pocosin habitats (Fig. 4). Our results from these sites were more limited in number, but they did provide insight into net vertical elevation processes occurring in these upslope systems. These systems provide important ecosystem functions today and provide the future horizontal transgression space that will allow marshes to thrive and migrate upriver and upslope today and into the future (Kirwan et al. 2016; Stagg et al. 2016; Osland et al. 2022b). Horizontal transgression has already been observed in the low-elevation forests at Alligator River NWR in North Carolina where more than 19,000 hectares of forested wetlands have transitioned to marsh or shrubland habitat in the past 35 years, likely due to saltwater intrusion from storms and SLR (Table S1; Ury et al. 2021). The transition of forested wetlands and pocosins to scrub-shrub and marsh habitat has been documented across the coastal plain (Ury et al. 2021; White et al. 2022). This has implications for carbon storage, carbon cycling, and ecosystem function (Aguilos et al. 2021). Strategic acquisition and conservation of conservation corridors upstream and upslope of present-day coastal wetlands is a critical component to conserving marshes, pocosins and forested wetlands in the future and has been recommended as a key conservation strategy (ACJV 2019).
Surface elevation at the forested wetland site at Roanoke River NWR, located at the mouth of the Roanoke River where it meets the Albemarle Sound, was gaining elevation at a highly variable rate (3.4 ± 0.8 mm/year; Table 3) that was less than the rate of SLR. Hence, this estuary is slowly inundating the islands at the lower end of the Roanoke River floodplain. This site had consistently wet soils that maintained integrity by supporting the anaerobic conditions that prevented the oxidation of peat soils and eliminated any detectable subsidence. Recent gains in surface elevation in this environment were likely attributed to belowground expansion, as no measurable accretion was observed at the site. Other studies of tidal freshwater forested wetlands in the South Atlantic geography have also found minimal accretion in these systems and have attributed wetland elevation gain to subsurface processes, such as root zone expansion (Noe et al. 2016; Stagg et al. 2021). Management strategies at Roanoke River NWR are now focusing on land acquisition to enable upslope wetland migration through the development of a wetland habitat expansion plan.
The pocosin sites (Alligator River — Pocosin and Pocosin Lakes NWRs) were losing elevation at a rate of approximately 9.0 mm/year. Previous studies have estimated subsidence to be as high as 20 mm/year in pocosin wetlands with altered hydrology (Richardson 1983). The subsidence rates we observed (15.0 mm/year at Alligator River and 9.5 mm/year at Pocosin Lakes NWRs, respectively) were less than half of those previously published, which is likely because we intentionally selected sites with minimal alterations to hydrology. Much of the pocosin habitat on the Albemarle-Pamlico Peninsula, North Carolina, was previously ditched and drained for conversion to agriculture. This has resulted in a lowering of the groundwater table, soil decomposition due to oxidation and saltwater intrusion, increased subsidence and inundation, and increased frequency of large, catastrophic peat fires (Faustini et al. 2020). Because of these combined synergistic effects, the volume of peat on the Albemarle-Pamlico Peninsula is approximately less than half of its historic volume (Lilly 1981). Alligator River NWR has already exhibited transgression of marshes into upslope forested and pocosin wetlands (Ury et al. 2021). Considering the site at Pocosin Lakes NWR had greater elevation capital and is located more inland than the Alligator River NWR site, we can assume a longer timeframe before upslope migration of marshes occurs. In all, the results from our study support the idea that pocosin restoration implemented to rewet peat soils for the purpose of storing carbon, reducing the risk of catastrophic fires and maintaining pocosin habitat for wildlife, particularly in areas with high elevation capital, should be prioritized (Faustini et al. 2020).
Implications for Management
In response to concerns surrounding coastal wetland ecosystems, USFWS has started to act through their Climate Change Action Program, which recommends the use of the resist-accept-direct (RAD) strategies for long-term adaptation planning associated with ecological transformation (Schuurman et al. 2020; USFWS 2021). This framework can be used to identify plausible future scenarios for a coastal wetland habitat, weigh the costs and benefits associated with various management strategies designed to resist, accept, or direct transformation, and then develop an overall portfolio of strategies to ensure the greatest conservation of coastal wetlands across the landscape. Considering one of the purposes of NWRs is to provide a place for learning, NWR coastal wetlands provide an ideal setting for applying RAD strategies to consider what management techniques will provide the best social, ecological, and economic outcomes. Blackwater NWR is an excellent example of a NWR using a mix of all three RAD strategies to develop their Blackwater 2100 plan (Schuurman et al. 2020).
Outside of NWRs, these findings are also applicable in minimally disturbed coastal marshes and can assist in the construction of effective conservation portfolios for public and privately owned coastal wetlands. In coastal wetlands, conservation and preservation strategies should focus on building vertical elevation capital, protecting horizontal transgression spaces upslope and upriver, or creating new habitats inland (ACJV 2019). Conservation of transgression spaces is a key component of these strategies. More specifically, the conservation of oligohaline marshes is critical, as they represent one of the key upriver transgression spaces for salt marshes in the future, currently have the potential to keep pace with SLR, and are predicted to experience net losses due to these transgressions (Osland et al. 2022b). In addition to oligohaline marshes, the conservation of forested wetland and pocosins is important, as they represent both an important habitat today and the future transgression space for upslope migration of coastal marshes. Strategies aimed at effectively directing transgressions that are already occurring and reducing subsidence will need to be considered as landscape conservation designs are developed and implemented (Faustini et al. 2020; Ury et al. 2021; Osland et al. 2022b). Finally, improving our understanding of the role of the tidal regime on the biophysical processes of coastal marshes and how each unique marsh system responds to restoration strategies, such as prescribed fire and thin-layer sediment deposition, will help us better restore and maintain habitats in the near term (McKee and Grace 2012; Powell et al. 2019; Faustini et al. 2020; Moon et al. 2022). An overall strategy that involves incorporating multiple RAD strategies across the NWR System and at the scale of an individual NWR will minimize the loss of coastal wetland habitats across the South Atlantic geography and the wildlife species dependent on them.
Data Availability
The data that support the findings of this study are openly available in USFWS's Service Catalog (ServCat) at https://ecos.fws.gov/ServCat/Reference/Profile/34452, reference number 34452.
References
Aguilos, M., C. Brown, K. Minick, M. Fischer, O.J. Ile, D. Hardesty, M. Kerrigan, A. Noormets, and J. King. 2021. Millennial-scale carbon storage in natural pine forests of the North Carolina lower coastal plain: Effects of artificial drainage in a time of rapid sea level rise. Land 10: 1294. https://doi.org/10.3390/land10121294.
Atlantic Coast Joint Venture (ACJV). 2019. Salt marsh bird conservation plan for the Atlantic coast. Atlantic Coast Joint Venture. https://conservationstandards.org/wp-content/uploads/sites/3/2020/10/salt_marsh_bird_plan_final_web.pdf. Accessed 11 January 2023.
Barbier, E.B., S.D. Hacker, C. Kennedy, E.W. Koch, A.C. Stier, and B.R. Silliman. 2011. The value of estuarine and coastal ecosystem services. Ecological Monographs 81: 169–193. https://doi.org/10.1890/10-1510.1.
Boyle, M.F., N.M. Rankin, and W.D. Stanton. 2015. Vegetation of coastal wetland elevation monitoring sites on National Wildlife Refuges in the South Atlantic geography: Baseline inventory report. U.S. Fish and Wildlife Service. https://ecos.fws.gov/ServCat/DownloadFile/108262?Reference=68327.
Cahoon, D.R., K.L. McKee, and J.T. Morris. 2021. How plants influence resilience of salt marsh and mangrove wetlands to sea-level rise. Estuaries and Coasts 44: 883–898. https://doi.org/10.1007/s12237-020-00834-w.
Cahoon, D.R., J.W. Day, D.J. Reed, and R.S. Young. 1998. Vulnerability of coastal wetlands in the Southeastern United States. In Global Climate Change and Sea-level Rise: Estimating the Potential for Submergence of Coastal Wetlands, ed. G.R. Guntenspergen and B.A. Vairin, 19–32. Reston, Virginia: Biological Resources Division, U.S. Geological Survey.
Cahoon, D.R., M.A. Ford, and P.F. Hensel. 2004. Ecogeomorphology of Spartina patens-dominated tidal marshes: Soil organic matter accumulation, marsh elevation dynamics, and disturbance. In The Ecogeomorphology of Tidal Marshes, ed. S. Fagherazzi, B. Marani and L.K. Blum, 247–266. Washington, DC: American Geophysical Union.
Cain, M.R., and P.F. Hensel. 2018. Wetland elevations at sub-centimeter precision: Exploring the use of digital barcode leveling for elevation monitoring. Estuaries and Coasts 41: 582–591. https://doi.org/10.1007/s12237-017-0282-6.
Comer, P.J., D. Faber-Langendon, R. Evans, S.C. Gawler, C. Josse, G. Kittel, S. Menard, et al. 2003. Ecological systems of the United States: A working classification of U.S. terrestrial systems. NatureServe. https://www.natureserve.org/sites/default/files/pcom_2003_ecol_systems_us.pdf. Accessed 11 January 2023.
Cook, C.E., M.F. Boyle, and N.M. Rankin. 2016. Vegetation of coastal wetland elevation monitoring sites on National Wildlife Refuges in the South Atlantic geography: 1st status assessment report. U.S. Fish and Wildlife Service. https://ecos.fws.gov/ServCat/DownloadFile/138965?Reference=91821.
Costanza, R., R. de Groot, P. Sutton, S. van der Ploeg, S.J. Anderson, I. Kubiszewski, S. Farber, and R.K. Turner. 2014. Changes in the global value of ecosystem services. Global Environmental Change 26: 152–158. https://doi.org/10.1016/j.gloenvcha.2014.04.002.
Covington, S. 2020. An inventory of surface elevation tables installed on National Wildlife Refuge System lands. U.S. Fish and Wildlife Service. https://ecos.fws.gov/ServCat/DownloadFile/169733?Reference=115052.
Craft, C. 2007. Freshwater input structures soil properties, vertical accretion, and nutrient accumulation of Georgia and U.S tidal marshes. Limnology and Oceanography 52: 1220–1230. https://doi.org/10.4319/lo.2007.52.3.1220.
Craft, C., J. Clough, J. Ehman, S. Joye, R. Park, S. Pennings, H. Guo, and M. Machmuller. 2009. Forecasting the effects of accelerated sea-level rise on tidal marsh ecosystem services. Frontiers in Ecology and the Environment 7: 73–78. https://doi.org/10.1890/070219.
Crotty, S.M., C. Ortals, T.M. Pettengill, L. Shi, M. Olabarrieta, M.A. Joyce, A.H. Altieri, et al. 2020. Sea-level rise and the emergence of a keystone grazer alter the geomorphic evolution and ecology of southeast US salt marshes. Proceedings of the National Academy of Sciences of the United States of America 117: 17891–17902. https://doi.org/10.1073/pnas.1917869117.
Daily, G., S. Alexander, P. Ehrlich, J. Lubchenco, P. Matson, H. Mooney, S. Postel, S. Schneider, and D. Tilman. 1997. Ecosystem services: Benefits supplied to human societies by natural ecosystems. Issues in Ecology 2: 1–16. https://www.esa.org/wp-content/uploads/2013/03/issue2.pdf.
Dangendorf, S., C. Hay, F. Calafat, M. Marcos, C. Piecuch, K. Berl, and J. Jensen. 2019. Persistent acceleration in global sea-level rise since the 1960s. Nature Climate Change 9: 705–710. https://doi.org/10.1038/s41558-019-0531-8.
Faustini, J., Moorman, M.C., and Gerlach, S. 2020. Water resource inventory and assessment: Pocosin Lakes National Wildlife Refuge. U.S. Fish and Wildlife Service. https://www.fws.gov/sites/default/files/documents/PocosinLakesWRIA_Final_06182020.pdf.
Ganju, N., Z. Defne, K. Ackerman, B. Couvillion, M. Moorman. 2023. Applying geospatial metrics to guide monitoring and restoration in tidal wetlands. North Carolina surface elevation table (SET) community of practice workshop.
Gundersen, G., D.R. Corbett, A. Long, M. Martinez, and M. Ardón. 2021. Long-term sediment, carbon, and nitrogen accumulation rates in coastal wetlands impacted by sea level rise. Estuaries and Coasts 44: 2142–2158. https://doi.org/10.1007/s12237-021-00928-z.
Kamrath, B.J.W., M.R. Burchell, N. Cormier, K. W. Krauss, and D.J. Johnson. 2019. The potential resiliency of a created tidal marsh to sea level rise. Transactions of the ASABE 62: 1567–1577. https://doi.org/10.13031/trans.13438.
Karegar, M.A., T.H. Dixon, and S.E. Engelhart. 2016. Subsidence along the Atlantic Coast of North America: Insights from GPS and late Holocene relative sea level data. Geophysical Research Letters 43: 3126–3133. https://doi.org/10.1002/2016GL068015.
Kirwan, M., and P. Megonigal. 2013. Tidal wetland stability in the face of human impacts and sea-level rise. Nature 504: 53–60. https://doi.org/10.1038/nature12856.
Kirwan, M.L., G.R. Guntenspergen, A. D’Alpaos, J.T. Morris, S.M. Mudd, and S. Temmerman. 2010. Limits on the adaptability of coastal marshes to rising sea level. Geophysical Research Letters 37. https://doi.org/10.1029/2010GL045489.
Kirwan, M.L., S. Temmerman, E.E. Skeehan, G.R. Guntenspergen, and S. Fagherazzi. 2016. Overestimation of marsh vulnerability to sea level rise. Nature Climate Change 6: 253–260. https://doi.org/10.1038/nclimate2909.
Krasting, J.P., J.P. Dunne, R.J. Stouffer, and R.W. Hallberg. 2016. Enhanced Atlantic sea-level rise relative to the Pacific under high carbon emission rates. Nature Geoscience 9: 210–214. https://doi.org/10.1038/ngeo2641.
Kuznetsova, A., P.B. Brockhoff, and R.H.B. Christensen. 2017. lmerTest Package: Tests in linear mixed effects models. Journal of Statistical Software 82: 1–26. https://doi.org/10.18637/jss.v082.i13.
Ladin, Z., and M. Moorman. 2021. USFWS, Southeast region surface elevation table and marker horizon data analysis. U.S. Fish and Wildlife Service. https://ecos.fws.gov/ServCat/DownloadFile/118219?Reference=76193.
Lagomasino, D., D.R. Corbett, and J.P. Walsh. 2013. Influence of wind-driven inundation and coastal geomorphology on sedimentation in two microtidal marshes, Pamlico River estuary, NC. Estuaries and Coasts 36: 1165–1180. https://doi.org/10.1007/s12237-013-9625-0.
Langston, A., C. Alexander, M. Alber, and M. Kirwan. 2021. Beyond 2100: Elevation capital disguises salt marsh vulnerability to sea-level rise in Georgia, USA. Estuarine, Coastal and Shelf science 249. https://doi.org/10.1016/j.ecss.2020.107093.
Lenth, R. 2022. ‘Emmeans: Estimated marginal means, aka least-square means, R package version 1.7.5. https://CRAN.R-project.org/package=emmeans. Accessed 30 December 2022.
Lilly, J.P. 1981. A history of swamp land development in North Carolina. In Pocosin wetlands: an integrated analysis of Coastal Plain freshwater bogs in North Carolina, ed. C.J. Richardson, 20–39. Stroudsburg, PA: Hutchinson Ross.
Lynch, J.C., P. Hensel, and D.R. Cahoon. 2015. The surface elevation table and marker horizon technique: A protocol for monitoring wetland elevation dynamics. National Park Service. https://irma.nps.gov/DataStore/DownloadFile/531681.
McKee, K.L., and J.B. Grace. 2012. Effects of prescribed burning on marsh-elevation change and the risk of wetland loss. U.S. Geological Survey Open-File Report 2012–1031: 51. https://doi.org/10.3133/ofr20121031.
Moon, J.A., L.C. Feher, T.C. Lane, W.C. Vervaeke, M.J. Osland, D.M. Head, B.C. Chivoiu, et al. 2022. Surface elevation change dynamics in coastal marshes along the Northwestern Gulf of Mexico: Anticipating effects of rising sea-level and intensifying hurricanes. Wetlands 42: 49. https://doi.org/10.1007/s13157-022-01565-3.
Moorman, M.C., and N. Rankin. 2020. South Atlantic-Gulf and Mississippi-Basin protocol framework for coastal wetland elevation monitoring. U.S. Fish and Wildlife Service. https://ecos.fws.gov/ServCat/DownloadFile/178825.
Moorman, M.C., J.M. McDonald, N.M. Rankin, and A.S. Mulligan. 2019. Determining elevation of rod surface elevation table benchmarks at coastal wetland elevation monitoring sites: Baseline report. U.S. Fish and Wildlife Service. https://ecos.fws.gov/ServCat/DownloadFile/168781.
Morris, J.T., P.V. Sundareshwar, C.T. Nietch, B. Kjerfve, and D.R. Cahoon. 2002. Responses of coastal wetlands to rising sea level. Ecology 83: 2869–2877. https://doi.org/10.1890/0012-9658(2002)083[2869:ROCWTR]2.0.CO;2.
National Oceanic and Atmospheric Administration (NOAA), Office for Coastal Management. 2016. Coastal change analysis program (C-CAP) regional land cover and change. Charleston, SC: NOAA Office for Coastal Management. www.coast.noaa.gov/htdata/raster1/landcover/bulkdownload/30m_lc/.
National Oceanic and Atmospheric Administration, Office for Coastal Management (NOAA). 2022. Sea level trends - NOAA tides & currents. https://tidesandcurrents.noaa.gov/sltrends/. Accessed 11 January 2023.
Noe, G.B., C.R. Hupp, C.E. Bernhardt, and K.W. Krauss. 2016. Contemporary deposition and long-term accumulation of sediment and nutrients by tidal freshwater forested wetlands impacted by sea level rise. Estuaries and Coasts 39: 1006–1019. https://doi.org/10.1007/s12237-016-0066-4.
Odum, W.E., E.P. Odum, and H.T. Odum. 1995. Nature’s pulsing paradigm. Estuaries 18: 547–555. https://doi.org/10.2307/1352375.
Osland, M.J., A.R. Hughes, A.R. Armitage, S.B. Scyphers, J. Cebrian, S.H. Swinea, C.C. Shepard, et al. 2022b. The impacts of mangrove range expansion on wetland ecosystem services in the southeastern United States: Current understanding, knowledge gaps, and emerging research needs. Global Change Biology 28: 3163–3187. https://doi.org/10.1111/gcb.16111.
Osland, M.J., B. Chivoiu, N.M. Enwright, K.M. Thorne, G.R. Guntenspergen, J.B. Grace, L.L. Dale, et al. 2022a. Migration and transformation of coastal wetlands in response to rising seas. Science Advances 8: eabo5174. https://doi.org/10.1126/sciadv.abo5174.
Palinkas, C.M., and K.A.M. Engelhardt. 2019. Influence of inundation and suspended-sediment concentrations on spatiotemporal sedimentation patterns in a tidal freshwater marsh. Wetlands 39: 507–520. https://doi.org/10.1007/s13157-018-1097-3.
Pickens, B.A., R.S. Mordecai, C.A. Drew, L.B. Alexander-Vaughn, A.S. Keister, H.L.C. Morris, and J.A. Collazo. 2017. Indicator-driven conservation planning across terrestrial, freshwater aquatic, and marine ecosystems of the South Atlantic, USA. Journal of Fish and Wildlife Management 8: 219–233. https://doi.org/10.3996/062016-JFWM-044.
Powell, E.J., M.C. Tyrrell, A. Milliken, J.M. Tirpak, and M.D. Staudinger. 2019. A review of coastal management approaches to support the integration of ecological and human community planning for climate change. Journal of Coastal Conservation 23: 1–18. https://doi.org/10.1007/s11852-018-0632-y.
R Core Team. 2022. R: A language and environment for statistical computing (version 4.2.0). R Foundation for Statistical Computing.
Reed, D.J. 2002. Sea-level rise and coastal marsh sustainability: Geological and ecological factors in the Mississippi delta plain. Geomorphology 48: 233–243.Richardson, C. J. 1983. Pocosins: Vanishing Wastelands or Valuable Wetlands? BioScience 33: 626–633. https://doi.org/10.2307/1309491.
Richardson, C.J. 1983. Pocosins: Vanishing wastelands or valuable wetlands? BioScience 33: 626–633. https://doi.org/10.2307/1309491.
Saba, V.S., S.M. Griffies, W.G. Anderson, M. Winton, M.A. Alexander, T.L. Delworth, J.A. Hare, et al. 2016. Enhanced warming of the Northwest Atlantic Ocean under climate change. Journal of Geophysical Research: Oceans 121: 118–132. https://doi.org/10.1002/2015JC011346.
Saintilan, N., K.E. Kovalenko, G. Guntenspergen, K. Rogers, J.C. Lynch, D.R. Cahoon, C.E. Lovelock, et al. 2022. Constraints on the adjustment of tidal marshes to accelerating sea level rise. Science 377: 523–527. https://doi.org/10.1126/science.abo7872.
Sallenger, A.H., K.S. Doran, and P.A. Howd. 2012. Hotspot of accelerated sea-level rise on the Atlantic coast of North America. Nature Climate Change 2: 884–888. https://doi.org/10.1038/nclimate1597.
Schuurman, G., H.-H. Cat, D. Cole, D. Lawrence, J. Morton, D. Magness, A. Cravens, S. Covington, R. O’Malley, and N. Fisichelli. 2020. Resist-accept-direct (RAD)—a framework for the 21st-century natural resource manager. National Park Service. https://doi.org/10.36967/nrr-2283597.
Siegert, M.R.B., E. Alley, J. Englander. Rignot, and R. Corell. 2020. Twenty-first century sea- level rise could exceed IPCC projections for strong-warmer futures. One Earth 3: 691–703. https://doi.org/10.1016/j.oneear.2020.11.002.
Silvestri, S., and M. Marani. 2004. Salt-marsh vegetation and morphology: Basic physiology, modelling, and remote sensing observations. In The Ecogeomorphology of Tidal Marshes, ed. S. Fagherazzi, B. Marani, and L.K. Blum, 5–25. Washington, DC: American Geophysical Union.
Stagg, C.L., K.W. Krauss, D.R. Cahoon, N. Cormier, W.H. Conner, and C.M. Swarzenski. 2016. Processes contributing to resilience of coastal wetlands to sea-level rise. Ecosystems 19: 1445–1459. https://doi.org/10.1007/s10021-016-0015-x.
Stagg, C.L., M.J. Osland, J.A. Moon, L.C. Feher, C. Laurenzano, T.C. Lane, W.R. Jones, and S.B. Hartley. 2021. Extreme precipitation and flooding contribute to sudden vegetation dieback in a coastal salt marsh. Plants 10: 1841. https://doi.org/10.3390/plants10091841.
Sweet, W.V., R. Kopp, C.P. Weaver, J. Obeysekera, R.M. Horton, E.R. Thieler, and C.E. Zervas. 2017. Global and regional sea level rise scenarios for the United States. National Oceanic and Atmospheric Administration, National Ocean Service, Center for Operational Oceanographic Products and Services. https://doi.org/10.7289/V5/TR-NOS-COOPS-083.
Sweet, W.V., B.D. Hamlington, R.E. Kopp, C.P. Weaver, P.L. Barnard, D. Bekart, W. Brooks, et al. 2022. Global and regional sea level rise scenarios for the United States: Updated mean projections and extreme water level probabilities along U.S. coastlines. National Ocean and Atmospheric Administration, National Ocean Service. https://oceanservice.noaa.gov/hazards/sealevelrise/noaa-nostechrpt01-global-regional-SLR-scenarios-US.pdf.
U.S. Fish and Wildlife Service (USFWS). 2021. Climate change action program summary. https://www.fws.gov/initiative/climate-change/climate-change-action-program.
U.S. Fish and Wildlife Service (USFWS). 2022. USFWS, Southeast region surface elevation table and marker horizon data analysis. https://ecos.fws.gov/ServCat/Reference/Profile/146614.
Ury, E.A., X. Yang, J.P. Wright, and E.S. Bernhardt. 2021. Rapid deforestation of a coastal landscape driven by sea‐level rise and extreme events. Ecological Applications 31. https://doi.org/10.1002/eap.2339.
Vogel, R.L., B. Kjerfve, and L.R. Gardner. 1996. Inorganic sediment budget for the North Inlet salt marsh, South Carolina, U.S.A. Mangroves and Salt Marshes 1: 23–35. https://doi.org/10.1023/A:1025990027312.
Wasson, K., N.K. Ganju, Z. Defne, C. Endris, T. Elsey-Quirk, K.M. Thorne, C.M. Freeman, G. Guntenspergen, D.J. Nowacki, and K.B. Raposa. 2019. Understanding tidal marsh trajectories: Evaluation of multiple indicators of marsh persistence. Environmental Research Letters 14: 124073. https://doi.org/10.1088/1748-9326/ab5a94.
White, E.E., E.A. Ury, E.S. Bernhardt, and X. Yang. 2022. Climate change driving widespread loss of coastal forested wetlands throughout the North American Coastal Plain. Ecosystems 25: 812–827. https://doi.org/10.1007/s10021-021-00686-w.
Yin, J. 2023. Rapid decadal acceleration of sea level rise along the U.S. east and gulf coasts during 2010 – 2022 and its impact on hurricane-induced storm surge. Journal of Climate (published online ahead of print 2023). https://doi.org/10.1175/JCLI-D-22-0670.1
Acknowledgements
We want to acknowledge the contributions of Ken Krauss, Joe DeVivo, Brian Boutin, Tony Curtis, Don Cahoon, Nicole Cormier, and Jim Lynch to the project’s conceptualization and study design when it was first implemented in 2011. We want to specifically thank David O’Loughlin and Jeremy Schmid of Atkins engineering for their help with SET installation, Kevin Keeler (USFWS retired) for leading monitoring of the SET at Cedar Island NWR for 8 years, Mike Chouinard (USFWS) for coordinating the program while it was in transition, and all additional personnel who have assisted with data collection for this project over the past 10 years. We would also like to extend our appreciation to the Associate Editor and three anonymous reviewers for their helpful comments and suggestions for this manuscript.
Funding
This research has been funded by the U.S. Fish and Wildlife Service southeast region’s Inventory and Monitoring Branch. The findings and conclusions in this article are those of the author(s) and do not necessarily represent the views of the U.S. Fish and Wildlife Service.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Charles T. Roman
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Moorman, M.C., Ladin, Z.S., Tsai, E. et al. Will They Stay or Will They Go — Understanding South Atlantic Coastal Wetland Transformation in Response to Sea-Level Rise. Estuaries and Coasts (2023). https://doi.org/10.1007/s12237-023-01225-7
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12237-023-01225-7